|
アヤメ属植物に含まれるイソフラボノイド成分とその活性
国立科学博物館 筑波実験植物園 岩科 司(日本花菖蒲協会 顧問)
1・イソフラボノイドとは
ハナショウブやアヤメ、カキツバタの花の赤紫〜紫の色は主にエンサチンやペタニンなどのアントシアニンに起因していることはこれまでに多く報告されている。このアントシアニンは広くはフラボノイドと総称される化合物群の1クラスである。またやはり花(および葉)に含まれており、上記のアントシアニンと細胞内でコピグメント効果を発現し、花色をより青味を帯びさせる働きのあるイソビテキシンやスウェルチジンのようなフラボンもまたフラボノイドの仲間である。このように一見してフラボノイド化合物はすべて色素として発現するように思われがちであるが、現在までに四千を越える種類が植物から報告されているフラボノイドの多くはまったく色を持たない。したがって色素としての機能はない。
フラボノイドは上述したアントシアニンやフラボン、それにフラボノール、フラバノン、カルコンなど、いくつかのクラスに分けることが出来るが、C6-C3-C6を基本骨格とするフラボノイドの中で、アントシアニンやフラボンがB-環と呼ばれるベンゼン環がC-環の2-位に結合しているのに対して、-位に結合しているフラボノイドをイソフラボノイドと呼んでいる(図1)。このイソフラボノイドにもまたいくつかの種類があり、イソフラボン、イソフラバノン、ロテノン、プテロカルパン、クマラノクロモンなどに細分される。これらのイソフラボノイドの中で、これまで最も多くの種類が報告されているのがイソフラボンで、一九九六年の段階で二八四種類が植物から分離されている。アントシアニンなどとは異なり、-環のベンゼン環が2-位から3-位に結合位置を換えただけで、物質の色素としての働きがまったく失われてしまうことは面白い。しかしその代わりに、他のフラボノイドには見られない多くの活性があらわれる。その事には後に触れるとして、イソフラボノイドはあらゆる植物から報告されているわけではなく、いくつかの植物に限って見出されている。今日までにキク科、クワ科、オシロイバナ科など双子葉植物15科、ショウガ科など単子葉植物4科、それに裸子植物2科(マキ科とヒノキ科)、蘚苔類1科(ハリガネゴケ科)から報告があるのみである。
その中で極端に分布の集中しているのがマメ科で、現在までに自然界で分離同定されたイソフラボノイドの実に9割以上がこの科で報告されている。そして、次に多く報告されているのがアヤメ科である。中でもアヤメ属では種類を越えるイソフラボノイドが分離されている。アヤメ科に限らず、この化合物群は時に、葉、心材、樹皮、花などからも見出されるが、ほとんどの場合は地下部、特に根や塊茎、それに種子で多く発生し、含有量も多い。
2・アヤメ属のイソフラボノイド
アヤメ属に含まれるイソフラボノイドの報告は古く、すでに十九世紀の後半にはニオイイリス(I. florentina)の塊茎からイリゲニン -グルコシド(イリジン)が分離同定されていた(De Laire and Tiemann, 1893)。現在までに種のアヤメ属植物から種類を越えるイソフラボノイドが報告されている(表1)。これらの物質の化学構造については2、3の代表例を除いてここでは詳細を省略する。後述の文献を参照されたい。これらの成分は他の植物と同様にそのほとんどが塊茎からの報告である。I. lutescensやI. pseudopumilaでは葉から分離されているが、これらの植物では地下部が分析されておらず、おそらく地下部にはより多量にイソフラボノイドが含まれていると推定される。
分離されたイソフラボノイドのほとんどはイソフラボンで、I. spuriaとI. croceaからそれぞれ、12a-ヒドロキシロテノンのイリスプリノール (irispurinol)と9-メトキシイリスプリノール (9-methoxyirisupurinol)が、またキショウブからクマラノクロモンのアヤメニンA〜E (ayamenin A〜E)、ルピナルビンA (lupinalbin A)および5,7,3´-トリヒドロキシ-6-メトキシクマラノクロモン(5,7,3-trihydroxy-6-methoxycoumaranochromone)が分離されているが、これらは塩化第二銅の処理によってストレスを与えられた葉から検出されたもので、正常な葉には存在しない。
アヤメ属植物から分離されたイソフラボノイドのほとんどはその物質名、例えばイリソン(irisone)、イリリン(irilin)、イリステクトリゲニン(iristectorigenin)、イリゲニン(irigenin)、イリロン(irilone)、イリスフロレンチン(irisflorentin)などの名前から推察されるように、アヤメ属固有の成分か、あるいはアヤメ属で始めて分離された成分であり、明らかにマメ科のものとは異なっている。
アヤメ属のイソフラボノイドは遊離の状態、すなわち糖が結合していないものが一般的であるが、7-グルコシドや4´-グルコシドなどの配糖体としても存在する。
アヤメ属植物から分離されたイソフラボノイドの特徴として、6,7-メチレンジオキシル化(イリスフロレンチン、イリフロゲニン、イリソロンなど)、6-あるいは8-位、あるいはその両方のヒドロキシル化またはメトキシル化(イリステクトリゲニン、テクトリゲニン、イリソリドンなど)、2´-位のヒドロキシル化またはメトキシル化(イリソンA、イリソンBなど)があげられる。また塩化第二銅での処理によってストレスを与えられたキショウブの葉で産生されるイリリンCとゲニステインを除いて、必ず一ヶ所あるいはそれ以上メトキシル化あるいはメチレンジオキシル化されている。これらは同一の植物の葉や花に含まれている-グリコシルフラボンやフラボノールともかなり異なった置換パターンであり、大多数は塊茎でその生合成が発現されている。しかしながら、メチレンジオキシル化や-位あるいは-位での置換の生合成経路は現在までわかっていない。
3・イソフラボノイドの 機能と活性
先にも述べたように、イソフラボノイドに属する成分は色素あるいはコピグメント物質としての働きがまったくない。他のフラボノイド、例えばアントシアニンやフラボン、フラボノールの場合、特に花に存在する場合は明らかに花粉媒介のための昆虫の誘引としての機能があるが、主として地下部に多く含まれるイソフラボノイドの機能については長い間不明であった。しかし、ダイズ、クローバー、クズなどのマメ科植物を中心に最近、その機能について若干明らかになりつつある。そのひとつはファイトアレキシン(phytoalexin)、すなわち正常な状態では存在しないが、病原菌に羅病されたときに菌を撃退するために植物が生産する活性物質、としての役割である (Ingham et al., 1983; Grayer and Harborne, 1994; 瓜谷・小島, 1977)。またその逆にマメ科のいくつかでは植物の生育に必要な根粒菌を根に誘引する物質がイソフラボノイドの一種であることも証明された。さらにアカクローバーなどでは連作障害を引き起こすのが自身の生産するイソフラボノイドであるとの報告もある (Tamura et al., 1967, 1969; Chang et al., 1969) 。
薬理および生理活性については、ダイズとその加工品に含まれるイソフラボンが抗菌、抗炎症、抗腫瘍など多くの活性を持つ事が示されて注目を浴びている以外に、人体に対する機能については枚挙にいとまがない。
現在までにアヤメ属植物から分離されたイソフラボノイドのうち、活性が認められているものを表2に示した。残念ながら、数種類がアヤメ属植物から分離されているにもかかわらず、その活性が証明されたのはわずか7種類のみで、その他の物質については活性の調査は行われていない。しかし、これらの中にはイリゲニン
7-グルコシドにみられるように、すでに実用的に用いられているものもある。また抗菌、抗酸化、抗腫瘍など重要な活性が認められているものもある。
今までにアヤメ属植物から分離されたイソフラボノイドのほとんどは構造中に必ず一つあるいはそれ以上のメトキシル基やメチレンジオキシル基をもつ。これらの基は一般に菌、時には他の植物に対しても毒性を示すと考えられているので、おそらくアヤメ属の塊茎において菌に対しての攻撃から自身を守るための機能をもつと推定される。
日本産のアヤメ属植物についてはハナショウブ、アヤメ、カキツバタ、ヒオウギアヤメ、ヒメシャガ、シャガなどいずれもイソフラボノイドについての報告がない。したがって、これらの植物の塊茎にイソフラボノイドがあるのかないのかは今日までまったく不明である。著者はこれまでアヤメ属植物の塊茎にイソフラボノイドが含まれており、アカクローバーにみられるようにハナショウブにおいても、これらの成分の自家中毒によって連作障害が発生するのではないかと推定していた。ところが私たちのグループで予備的にハナショウブ(金剛城)の塊茎を分析したところ、イソフラボノイドの性質を示す成分は検出されなかった(しかし、他の植物の芽生えを阻害する未知の成分は存在することが認められた)。もしかすると逆にハナショウブの場合、イソフラボノイドのような抗菌性を示す成分が欠落しているが故に、菌によって連作障害が発生するのかもしれない。あるいはその他の原因によるのか現在分析検討中である。
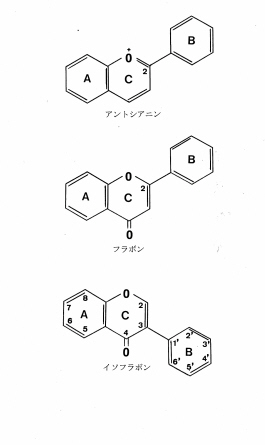 
図1 アントシアニン,フラボン及びイソフラボンの基本骨格図
表1 アヤメ属植物からのイソフラボノイドの報告
I. corcea
テクトリゲニン、テクトリゲニン - グルコシド、テクトリゲニン 4-グルコ シド、5,7-ジヒドロキシ-6,2-ジメトキ シイソフラボン、9-メトキシイリスプ リノール(塊茎)
(Shawl and Kumar, 1992)
ニオイイリス(I. florentina)
イリロン 4-グルコシド、イリソロン 4 ´-ビオシド(塊茎)(Tsukida et
al., 1973)、イリゲニン(塊茎)(柴田ら , 1960; Farkas and Varady,
1960; Morita et al., 1973; Arisawa et al., 1973a)、イリソロン、イリステクトリ ゲニンB、イリスフロレンチン(塊茎) (
Morita et al., 1973; Arisawa et al., 1973a)、イリゲニン 7-グルコシド(塊
茎)(De Laire and Tiemann, 1893; Baker, 1928; 柴田ら, 1960; Farkas
et al., 1966; Arisawa et al., 1973a)、イリ フロゲニン 4´-グルコシド(塊茎) (Arisawa
et al., 1973a)
ジャーマンアイリス(I. germanica)
イリゲニン(塊茎)(Bate-Smith and Swain, 1965; Pailer and Franke, 1973; El-Moghazy et al., 1980; Crawford et al., 1994)、イリゲニン 7-グルコシド (塊茎)(Dhar and Kalla, 1973; El- Moghazy et al., 1980)、イリロン(塊 茎)(Dhar and Kalla, 1973)、 テクトリゲニン 7-グルコシド、ホモテ クトリゲニン 7-グルコシド(塊茎) (Kawase et al., 1973)、テクトリゲニ ン(葉、塊茎)(Pailer and Franke, 1973; Williams et al., 1997)、 イリソロン*、2-ヒドロキシイリゲニ ン*(塊茎)(Crawford et al., 1994)、イ リソリドン、イリソロン、イリスフロ レンチン、5,3´,4´-トリメトキシ- 6,7-メチレンジオキシイソフラボン、 5,4´-ジヒドロキシ-6,7-メチレンジオ キシイソフラボン、5,7,3´-トリヒド ロキシ-6,4´-ジメトキシイソフラボ ン、イリステクトリゲニンB(塊茎) (Pailer and Franke, 1973)
I. germanica var. alba
イリソリドン7-グルコシド、5,3´-ジ ヒドロキシ-4´,5´-ジメトキシ-6,7- メチレンジオキシイソフラボン、5,7,3 ´-トリヒドロキシ-4´,5´-ジメト キシイソフラボン7-グルコシド(塊 茎)(Ali et al., 1983)
I. hookeriana
イリゲニン、イリゲニン 7-グルコシ ド、イリスフロレンチン、ユニペゲニ ンA(塊茎)(Shawl et al., 1985)
I. kashmiriana
イリスカシュミリアニン、イソイリス カシュミリアニン(塊茎)(Kachroo et al., 1990)、イリソリドン(塊茎)(Dhar and Kalla, 1975) I. kumaonensis
イリゲニン 7-グルコシド(全草、塊茎) (Dhar and Kalla, 1972; Agarwal et
al., 1984b)、イリスクマオニン(全草、塊 茎)(Kalla et al., 1978; Agarwal
et al., 1984b)、イリゲニン、イリスクマオニ ン メチル エーテル、イリスフロレン チン、ユニペゲニンA(塊茎)(Agarwal
et al., 1984b)
I. lutescens
テクトリゲニン(葉)(Williams et al., 1997)
I. milesii
プルネチン、テクトリゲニン、テクト リゲニン7-グルコシド、イリゲニン、 イリゲニン 7-グルコシド、5,6,7,4´- テトラヒドロキシ-8-メトキシイソフ ラボン、イリステクトリゲニンB 7- ま たは4´-グルコシド(塊茎)(Agarwal et al., 1984a)、5,6,7,4´-テトラヒドロ キシ-3´-メトキシイソフラボン(塊 茎)(Agarwal et al., 1984b)
I. missouriensis
イリソン A、イリソン B、5,7-ジヒド ロキシ-6,2´-ジメトキシイソフラボ ン(塊茎)(Wong
et al., 1987)
I. nepalensis
イリソロン(塊茎)(Gopinath et al., 1961; Prakash et al., 1965;
Bate-Smith and Swain, 1965; Fukui and Matsumoto, 1965; Ingham,
1983; Crawford et al., 1994)、 イリソリドン(塊茎)(Prakash et al.,
1965; Ingham, 1983)、イリゲニン(塊 茎) (Gopinath et al., 1963;
Prakash et al., 1965; Ingham, 1983)
I. nigricans
ニグリシン、ニグリカニン、イリロン、 5,3´-ジヒドロキシ-6,7-メチレンジオ キシイソフラボン(塊茎)(Al-Khalil et al., 1994)
I. pallida
イリゲニン7-グルコシド(塊茎)(Baker , 1928; Dean, 1963; Ingham, 1983)
キショウブ(I. pseudacorus) イリリン A**、イリリン B**、イリリ ン C**、イリステクトリゲニン
A**、 テクトリゲニン**、オロボール 3´- メチル エーテル**、プラテンセイン **、ビオカニン
A**、 ゲニステイン**、アヤメニン A**、ア ヤメニン B**、アヤメニン C**、アヤ メニン
D**、 5,7,3´-トリヒドロキシ-6-メトキシ クマラノクロモン**、ルピナルビン
A**(葉) (Hanawa et al., 1991a)、アヤメニン E**(葉)(Hanawa et al.,
1991b)
I. pseudopumila
テクトリゲニン(葉)(Williams et al., 1997)
I. spuria
イリステクトリゲニン A、イリステク トリゲニン A 7-グルコシド、5,7-ジヒ ドロキシ-6,2´-ジメトキシイソフラ ボン(塊茎)(Shawl
et al., 1984)、5,2 ´-ジヒドロキシ-7,8-ジメトキシイソ フラボン(塊茎)(Shawl
et al., 1988a)、 イリスプリノール(塊茎)(Shawl et al., 1988b)
イチハツ(I. tectorum)
テクトリゲニン(塊茎)(Bate-Smith and Swain, 1965)、テクトリゲニン
7- グルコシド(塊茎)(柴田, 1927; 朝比奈 ら, 1928; Seshadri, 1962)、イリゲニ ン7-グルコシド(塊茎)(Seshadri,
1962)、イリステクトリゲニン B 7-ま たは 4´-グルコシド(塊茎)(森田ら,
1972)、イリステクトリゲニン A 7-グ ルコシド(塊茎)(Morita et al., 1972)
I. tingitana
5,4´-ジメトキシ-6,7-メチレンジオキ シイソフラボン、3´-ヒドロキシ-5,4 ´-ジメトキシ-6,7-メチレンジオキシ イソフラボン(塊茎)(El-Emary et al., 1980)、イリゲニン、イリスフロレンチ ン(塊茎)(Ingham, 1983)
カンザキアヤメ(I. unguicularis)
イリゲニン、イリステクトリゲニン A (塊茎)(Arisawa and Morita, 1976;
Arisawa et al., 1976)、イリゲニン7- グルコシド(塊茎)(Arisawa et
al., 1976)
I. variegata
テクトリゲニン(葉)(Williams et al., 1997)
* 嫌気的条件で処理された植物でのみ生産
**塩化第二銅で処理された植物でのみ生産
表2 アヤメ属植物に含まれるイソフラボノイドの生理および薬理活性
イリロン
小麦の子葉鞘の成長を抑制
イリステクトリゲニンA
高血圧の抑制、カテコール O-メチルトランスフェラーゼおよびDOPAデカルボキシラーゼの阻害
テクトリゲニン
抗菌、ファイトアレキシン
ビオカニンA
エストロゲン、脂質およびコレステ ロールの低下
ゲニステイン
抗酸化、抗腫瘍、ダイズのリパーゼおよびパーエステラーゼ活性の阻害
イリゲニン
RNA合成の活性化
イリゲニン7-グルコシド
化粧品に用いられるニオイイリス塊茎 の主要成分
(引用文献は省略)
|