|
 ハナショウブにおける赤色花の育種戦略 ハナショウブにおける赤色花の育種戦略
宮崎大学農学部 藪谷 勤
1. はじめに
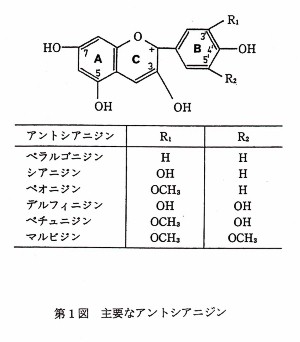
ハナショウブにおける多彩な花色の発現には、アントシアニンというフラボノイド色素が中心的な役割を果たしている。この色素は、花色を発現するもとのアントシアニジンに糖が結合した配糖体のことであり、時にはこの配糖体がクマール酸、カフェ酸などの有機酸によりアシル化された構造をとることもある。主要なアントシアニジンは第1図に示した6種類であるが、これらのアントシアニジンはB環の水酸化とメチル化の程度が異なるだけである。第1図に示したB環の水酸基が増加するにつれて赤色から青味がかっていく性質があり、ペラルゴニジンはオレンジ赤、シアニジンは赤、ペオニジンはマゼンタ(紫赤)、デルフィニジンは青、ペチュニジンおよびマルビジンは紫の花色を発現するのに重要な色素である。これらのアントシアニジンの内、ハナショウブではペラルゴニジンを除いた5種類が確認されている。
ハナショウブにおける花色の多彩化育種を促進するために、我々は花弁(外花被)含有アントシアニンに関する高速液体クロマトグラフィー(HPLC)を行い、その有用変異の発見に努めている。最近、赤色花を育種するのに極めて重要な色素である
シアニジン3RGやシアニジン3RG5Gを主要アントシアニンとする品種を見つけたので、本稿ではこれらのアントシアニンによる赤色花の育種戦略について述べたい。
2. 主要アントシアニンの変異
第1表は、ハナショウブにおいてこれまでに発見されている15種類の主要アントシアニンの型を示したものである。これらの型の内、マルビジン3RGac5G
- ペチュニジン3RGac5Gは本種における主要アントシアニンの基本(野生型)とみなされている(Yabuya
1991、Yabuya et al. 1994)。従って、その他の14種類の型はいずれも変異型であり、これらの内デルフィニジン3RGac5Gは青色花を発現するのに鍵となるアントシアニンである。このアントシアニンによる青色花の育種戦略については既に本誌(第25号)で述べた(藪谷
1997)。一方、ハナショウブには赤色系アントシアニンとしてシアニジン
3RGac5G、シアニジン 3RG5G、シアニジン3RG、ペオニジン
3RGac5G、ペオニジン3RG5Gおよびペオニジン3RGが主要色素として存在するにもかかわらず、赤色花品種が育成されるまでには至っていないのが現状である(第1、2表)。
 |
 |
3. 赤色花の育種戦略
ハナショウブにおける赤色系アントシアニンの主要成分の型と花色の関係を第2表に示した。これまでに育成されている品種の中では、「火の舞」(第2図A)や「火渡り」などが最も赤い品種であるが、いずれもペオニジン
3RGac5G型に属する(第2表)。しかしながら、これらの品種は赤色花というよりはマゼンタ花を発現しており、この色は先に述べたようにペオニジンから期待される花色でもある。これに対して、シアニジンおよびペラルゴニジンからは赤色花が期待されるが、ハナショウブのシアニジン
3RGac5G、シアニジン 3RGac5G - ペオニジン
3RGac5G、ペオニジン3RG5G - シアニジン
3RG5Gおよびペオニジン3RG
- シアニジン3RG型に属する品種(系統)はいずれもピンク系の花色しか発現していない(第2表、第2図B〜D)。
ペチュニアの赤色花はシアニジン 3Gおよびシアニジン3RGによることが報告されている(Wiering
and de Vlaming 1984)。ハナショウブではこれらアントシアニンの単独型品種は未だ発見されていないが、シアニジン
3RGac5Gの単独型品種(系統)はいずれもピンクの花色を有している(第2表)。また、ペチュニアでもシアニジン
3RGac5Gはマゼンタ花を発現している(Wiering
and de Vlaming 1984)。従って、ハナショウブに赤色花を育成するためには、シアニジン
3RGac5Gの脱アシル型、すなわちシアニジン
3RG5G、シアニジン3RGおよびシアニジン
3Gの単独型の獲得が必要である。これらの内、シアニジン
3RG5Gおよびシアニジン3RGの両単独型の獲得はペオニジン3RG5G
- シアニジン 3RG5Gまたはペオニジン3RG
- シアニジン3RG型の品種とシアニジン
3RGac5G型品種(系統)を交雑したF2個体(シアニジン
3RG5G がF2の約6.3%、シアニジン3RGがF2の約1.6%)に期待される。何故ならば、シアニジンのメチル化(ペオニジン)、アシル化(ac)および配糖化(5G)の各遺伝子に関する劣性ホモの個体がF2で得られるからである。このようにして獲得したシアニジン
3RG5Gやシアニジン3RG型の個体を材料にして交雑育種を行えば、赤色花品種の育成が可能であろう。
一方、ペラルゴニジン系アントシアニンにより赤色(オレンジ赤)花を育成することも考えられるが、このアントシアニンはハナショウブばかりでなく他のアヤメ(Iris)属植物においてもその存在は確認されていない。従って、ハナショウブにペラルゴニジン系アントシアニンによる赤色花品種を育成するためには、ペチュニア(Meyer
et al. 1987、Tanaka et al. 1995)においてこのアントシアニンの発現に成功した形質転換技術の導入が必要である。
4. 引用文献
Meyer, P., I. Heidmann, G. Forkmann
and H.
Saedler 1987. A new petunia flower
colour
generated by transformation of a mutant
with
a maize gene. Nature 330:677-678.
Tanaka, K., Y. Fukui, M. Fukuchi-Mizutani,
T. M. Holton, E. Higgins and T. Kusumi
1995.
Molecular cloning and characterization
of
Rosa hybrida dihydroflavonol 4-reductase
gene. Plant Cell Physiol.36:1023-1031.
Wiering, H. and P. de Vlaming 1984.
Inheritance
and biochemistry of pigments. p.49-67,
In:
K. C. Sink(Ed), Petunia.
Yabuya, T. 1991. High-performance liquid
chromatographic analysis of anthocyanins
in Japanese garden iris and its wild
forms.
Euphytica 52:215-219.
Yabuya, T., M. Nakamura and A. Yamasaki
1994.
P-coumaroyl glycosides of cyanidin
and peonidin
in the flowers of Japanese garden iris,
Iris
ensata Thunb. Euphytica 74:47-50. 藪谷 勤 1997.
ハナショウブにおける青色花の育種戦略。花菖蒲25:2-6。

|